(Z creation.com/biogeography přeložil M. T. – 02/2014. Tento článek je založen na 6. kapitole knihy D. Stathama Evolution: Good Science? /Evoluce: Dobrá věda?/)
Evolucionisté tvrdí, že biogeografické rozdělení organizmů svědčí přesvědčivě o evoluci. Třebaže biogeografické studie přinášejí pádné důkazy pro proces speciace, nejsou v souladu se širšími předpověďmi evoluční teorie ani nejsou v souladu s modelem pomalého kontinentálního driftu prosazovaným geology staré Země. Evoluční teorie má potíže s vysvětlením oblastí endemizmu i nespojitého rozmístění, které pozorujeme jak ve zkamenělém záznamu, tak v živé přírodě. Známá data tak svědčí pro biblickou zprávu o rekolonizaci po Potopě popsané v Genesis, a zejména pro hypotézu o tom, že pozorované vzorce vznikly z globálního rozmístění na přirozených raftech.

Obrázek 1. Placentální savci a jejich vačnatí protějšci.
Biogeografie je studium rozdělení rostlin a zvířat po světě. Z ní víme, že každý ze světadílů má vlastní odlišnou faunu a flóru. V Africe například nacházíme nosorožce, hrochy, lvy, hyeny, žirafy, zebry, šimpanze a gorily. V Jižní Americe žádné z těchto zvířat nežije. Místo toho jsou tam domovem pumy, jaguáři, mývalové, vačice a pásovci. Vačnatce nalézáme v Austrálii a Jižní Americe, nikoli však v Evropě. Taková pozorování vedla biogeografy k rozdělení světa na šest hlavních faunistických areálů. Podobně bylo zavedeno i šest hlavních floristických areálů. Evolucionisté tvrdí, že nejrozumnějším vysvětlením pro toto biogeografické rozrůznění je, že se ona odlišná zvířata a rostliny vyvinuly odděleně, z předků, kteří kolonizovali různé části světa před tisíci či miliony let. Další důkazy pro toto tvrzení prý plynou ze studia ostrovní biogeografie. Například z 1500 známých druhů octomilek (Drosophila) žije skoro třetina pouze na Havajských ostrovech. Tyto ostrovy jsou rovněž domovem více než 1000 druhů plžů a dalších suchozemských měkkýšů, které nenajdeme nikde jinde.
Zde je opět třeba rozlišovat mezi speciací v rámci druhu (která je přijímána jako fakt jak kreacionisty, tak evolucionisty) a evolucí mezi druhy (=druh je zde „kind“, ne „species“ – pozn. edit.). Biogeografie pak skutečně poskytuje důkazy ve prospěch prvé možnosti a zmíněné octomilky, plži i další měkkýši vyskytující se na Havajských ostrovech prokazatelně poskytují jedny z nejpádnějších důkazů takové speciace. Podobně existují jasné biogeografické důkazy pro speciaci pěnkav na galapážském souostroví, kde na jednotlivých ostrovech nalézáme podobné, ale ne zcela totožné druhy1. K tomu došlo téměř jistě proto, že ostrovy leží dost blízko na to, aby to umožnilo několika ptákům přeletět na sousední ostrov, ale dost daleko na to, aby byla nová ptačí kolonie izolována od původní skupiny a bylo tak méně pravděpodobné, že bude docházet k příbuzenskému křížení. Ale s jakým úspěchem se evoluční teorie vypořádává s obecnějšími biogeografickými pozorováními?
Ve skutečnosti se některá biogeografická pozorování mimořádně obtížně vysvětlují v rámci obrazu světa raženého evolucionisty. Podle evoluční teorie se savci vyvinuli zhruba před 100 miliony let z malých tvorů podobných rejskům. Z těchto tvorů se prý vyvinuli kromě jiného vačnatci žijící v Austrálii a placentálové žijící v Evropě i jinde ve světě. To, co je tak pozoruhodného na těchto dvou skupinách, je fakt, že i když jsou jejich rozmnožovací orgány zásadně odlišné, v jiných směrech jsou si tito tvorové velmi podobní (obrázek 1). Například kosterní struktury některých evropských placentálních psovitých šelem jsou téměř totožné se strukturami australských vačnatců-vakovlků. Zvlášť dobře je to patrné, srovnáme-li lebky bývalého tasmánského vakovlka (Thylacinus cynocephalus) a evropského placentálního vlka obecného (Canis lupus). Další placentálové a vačnatci, kteří se prý vyvíjeli nezávisle na sobě, vykazují rovněž podobné rysy. Lze opravdu věřit tomu, že se v důsledku náhodných mutací a rozdílného životního prostředí na oddělených světadílech vyvinuly u obou skupin tak podobné znaky?
(Kdo by ovšem chtěl vakovlka vidět, musel by patrně do muzea, neboť v přírodě ani v zajetí už zřejmě nežije. Prokazatelně poslední jedinec uhynul v roce 1936 v malé zoo v přístavu Hobart na Tasmánii. Od té doby sice občas proskočí zprávy o jeho pozorování, dokonce prý byly nalezeny i stopy a trus, avšak spolehlivý důkaz, že vakovlk ještě žije, chybí. Neuspěl ani známý horolezec E. Hillary, když po něm v 60. letech 20. století pátral v hlubokých lesích tasmánského vnitrozemí. Pozn. překl.)
Oblasti endemizmu
Jelikož se o evoluci tvrdí, že je celosvětovým jevem, očekávali bychom, že nové druhy budou vznikat na mnoha místech napříč všemi světadíly. Předpovědi evoluční teorie by tedy měly vycházet z toho, že střediska, odkud se šíří rostliny a zvířata, budou nepravidelně rozeseta, a nikoli soustředěna do několika málo oblastí2. Mnoho let už je však známo, že tomu tak není. Už roku 1820 si Augustin de Candolle uvědomil, že globální mapa míst původu rostlin se spíše blíží mapě „oblastí endemizmu“, kdy je mnoho různých rostlin omezeno vždy na jednu přesně vymezenou a často malou oblast (viz obrázek 4) 3. Následně se přišlo i na to, že se de Candollovy oblasti intenzivního rostlinného endemizmu kryjí rovněž s oblastmi intenzivního endemizmu živočišného4.
(Augustin Pyrame de Candolle, 1778-1841, švýcarský botanik; vytvořil teorii přirozeného systému rostlin, která nahradila umělou soustavu Linnéovu. Pozn. překl.)
Nespojité areály

Obrázek 2. Rozšíření rostliny rodu Clethra – jochovec (z Thorne, odkaz 9).
Dalším problémem, který evoluční biogeografie neumí vysvětlit, je fakt, že podobné rostliny a zvířata nalézáme nejen v navzájem navazujících oblastech souše či na sousedících ostrovech, nýbrž i na různých světadílech oddělených rozsáhlými plochami krajiny či oceánu. Takový výskyt nazýváme výskytem v nespojitých areálech. Evolucionisté tuto skutečnost někdy vysvětlují tak, že stěhování světadílů (kontinentální drift) rozdělilo podobné skupiny, které kdysi žily v těsné blízkosti, a měly tedy společné předky. Toto je vysvětlení uváděné například pro fakt, že některé pakomáry najdeme v Antarktidě, jižní Austrálii, Jižní Americe, na Novém Zélandu i v jižní Africe5. Avšak podle teorií samotných evolucionistů se mnoho druhů žijících odděleně na dříve spojených světadílech vyvinulo až po jejich rozdělení6. Například Jižní Amerika a Afrika se údajně oddělily zhruba před 100 miliony let, ale ty druhy kaktusů, které se údajně vyvinuly v Jižní Americe před zhruba třiceti miliony let, nalézáme i v Africe. Podobně evolucionistické výklady zrodu hlodavců žijících v Jižní Americe i Africe odporují obecně přijímanému datování kontinentálního driftu7. A je známo mnoho dalších zarážejících nespojitých areálů v obou těchto světadílech8. Navíc nalézáme druhy s nespojitými areály často na světadílech, které spolu nikdy nesousedily. Například je známo, že mnoho rostlin i hmyzu má nespojité areály v oblasti Tichého oceánu9. Rozšíření rostliny rodu Clethra – jochovec je dalším příkladem, znázorněným na obrázku 2. Zajímavé rovněž je, že vačnatec kolokolo (Dromiciops) žijící v Chile je mnohem příbuznější australským vačnatcům než jiným jihoamerickým vačnatcům10.
(Na tuto skutečnost ostatně upozorňuje i evolucionistický „Svět zvířat“, Albatros Praha 1997: „V poslední době totiž přichází stále více důkazů o tom, že kolokolo může být hledaným spojovacím článkem mezi jihoamerickými a australskými vačnatci. Nasvědčuje tomu i skutečnost, že přinejmenším šest vymřelých druhů obývalo před 20 miliony lety nejjižnější oblasti Jižní Ameriky.“ Pozn. překl.)
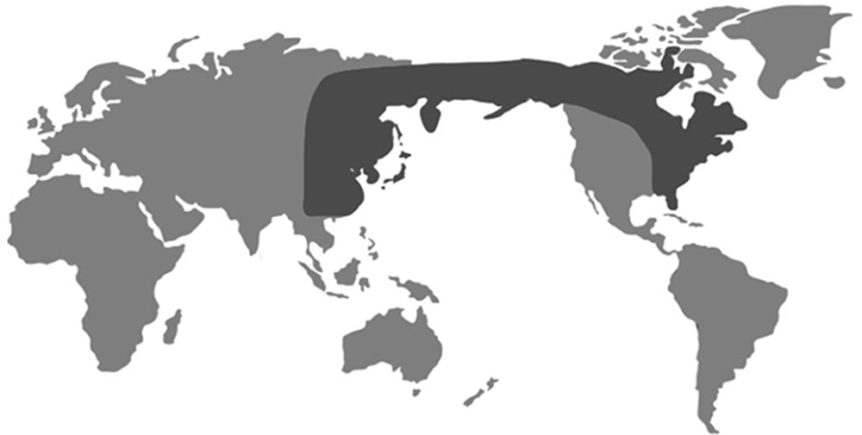
Lze uvést ještě spoustu dalších biogeografických anomálií, které nezapadají do očekávaného evolučního vzorce. Například fauna střední a jižní Afriky má blíže k fauně jižní Asie než k fauně severoafrické (11) a flóra nalézaná na Madagaskaru se neobyčejně podobá flóře indonéské12. Šichy (Empetrum) nalézáme pouze v severněji položených oblastech severní polokoule a v nejjižnějších oblastech jižní polokoule. Mnoho blízce příbuzných rostlin nacházíme pouze na východě Severní Ameriky a ve východní Asii. Ze studie provedené Muzeem státu Illinois vyplynulo, že ve východní Asii a na východě Severní Ameriky roste 627 rodů cévnatých rostlin, z nichž 151 chybí na západě Severní Ameriky13. Důležité je, že některé z rostlin (a hub) žijící ve východní Asii a na východě Severní Ameriky jsou totožné na úrovni druhů, což svědčí o tom, že k odloučení těchto areálů došlo teprve nedávno (to znamená během uplynulých pár tisíc let). Kdyby totiž k onomu odloučení došlo před mnoha miliony let, jak tvrdí evolucionisté, je velmi nepravděpodobné, že tolik druhů by v obou oblastech zůstalo stejných. Neboť je známo, že se rostliny i zvířata rychle mění v reakci na změny svého životního prostředí.
Fosilie
Fosilní záznam přináší rovněž problémy pro evoluční interpretaci biogeografie. Například na západě Severní Ameriky a ve východní Asii existuje mnoho podobných fosilií rostlin, ale podle teorie kontinentálního driftu preferované geology byly tyto horniny uloženy teprve poté, kdy už Aljašku a Rusko oddělovaly tisíce kilometrů oceánu14. Zatímco oblast výskytu žijících vačnatců tvoří téměř výhradně Austrálie a Jižní Amerika, jejich fosilie z období, které evolucionisté nazývají „svrchní křída“ (údajně před 65-85 miliony let) nalézáme výhradně v Eurasii a Severní Americe. Jak se vyjádřil Richard Cifelli, asistent na katedře zoologie oklahomské univerzity, „tento zeměpisný náhlý přesun zůstává nevysvětlen“15. Udivující je, že dnes nalézáme fosilní vačnatce na všech světadílech16. Podle evoluční teorie se placentálové vyvinuli na severní polokouli a v Austrálii se objevili teprve před zhruba pěti miliony let. Avšak nedávný objev čehosi, co se zdá být placentální fosilií, v Austrálii, v horninách starých údajně 120 milionů let, přiměl evolucionisty k vyslovení domněnky, že se placentálové zřejmě vyvinuli nejdřív na jižní polokouli, migrovali na sever, a pak asi na jižních světadílech vymřeli!17 O lvech se ví, že žili v Izraeli, ale lví fosilie se tam nenašly. Podobně se kdysi po USA proháněly miliony bizonů, ale nalézá se tam velmi málo fosilií bizonů. Tvrdit, že se určité zvíře s jistotou vyvinulo na určitém místě prostě proto, že se (dosud) nenašly důkazy o tom, že žilo někde jinde, není nutně vědecké.
Z těchto důvodů je jasné, že pozorované rozdělení organizmů nelze vysvětlit pouze poukazem na to, že se vyvinuly na místech, kde je nyní nacházíme. V důsledku toho rozšířili evolucionisté své modely biogeografie alternativními teoriemi jako třeba tvrzeními o migraci přes dříve existující mezikontinentální šíje, o dopravě pomocí ptáků a větru, a transoceánském šíření rostlin a zvířat na kobercích plovoucí vegetace18. V některých případech se tvrdí, že organizmy vyskytující se nyní v nespojitých areálech žily kdysi na souvislých územích a že takové rostliny či zvířata na územích zmíněné areály spojujících vymřely. Jinou teorií uváděnou pro vysvětlení zarážejících biogeografických pozorování je „konvergentní evoluce“. Podle ní se odlišné organizmy vyvinuly na různých místech světa v organizmy podobné, protože se musely přizpůsobit podobným biotopům. Tohle vysvětlení nabízejí evolucionisté například pro podobnosti mezi placentály a vačnatci19.
Při jakékoli diskuzi o biogeografických vzorcích bychom si měli uvědomit, že mnohé teorie, které se zde nabízejí, jsou nutně chudé na data, a v důsledku toho bohaté na fantazírování. Ke všem zkoumaným jevům totiž došlo mnoho let před okamžiky, ke kterým sahá lidská paměť, a mnoho z důkazů, které by snad podpořily ten který názor, zřejmě už dávno zmizelo. Možná je příznačné, že v devatenáctém století stavěli zastánci evoluční interpretace biogeografie na víře v oddělené, jasně dané světadíly, zatímco nyní se tvrdí, že pozorované vzorce života podporující evoluční výklad biogeografie počítají s kontinentálním driftem. Možná tedy je blíže pravdě názor vyjádřený Dr Garethem Nelsonem a Dr Normanem Platnickem z Amerického přírodovědného muzea, kteří tvrdí, že „se ukázalo, že biogeografie (čili zeměpisné rozšíření organizmů) nepřináší v žádném ohledu důkazy ani pro ani proti evoluci“20.
Avšak kreacionisté se mohou obrátit na Bibli jako na vodítko při snaze o porozumění celosvětovému rozšíření zvířat i rostlin. Podle ní začala rekolonizace světa bezprostředně po Potopě podle Genesis, když vody opadly (Genesis 8/. Zvířata vystoupila z archy a vegetace unášená proudy, nesoucí semena, hmyz i sladkovodní ryby, se uchycovala na zemi vystupující z vody. Kreacionistické modely se soustřeďují na čtyři hlavní procesy, které podle nich ovlivnily popotopní biogeografii:
- transoceánský transport na rostlinných kobercích
- transport obstaraný člověkem
- migrace a částečné vymření
- speciace.
Transoceánský transport na vegetačních kobercích
Už mnoho let přijímají evolucionisté i kreacionisté myšlenku možné přepravy rostlin i zvířat přes velké vodní plochy prostřednictvím přírodních koberců – naplavenin kmenů a keřů. Profesor Paul Moody z univerzity ve Vermontu prohlásil,
„V dobách záplav se mohou z říčních břehů odtrhnout velké kusy zeminy spolu s hustou vegetací včetně stromů a být smeteny do moře. Někdy lze zahlédnout takové ostrůvky-koberce na hladině oceánu daleko od pevniny, stále svěží a zelené, s palmami sedm až deset metrů vysokými. Je docela dobře možné, že tak mohou být přepravována na velké vzdálenosti i suchozemská zvířata. Mayr uvádí, že mnoho tropických oceánských proudů má rychlost přinejmenším dva uzle; to by činilo až osmdesát kilometrů za den, 1600 km za tři týdny“21.

Obrázek 3. Trasa, kterou urazil Oreobolus (z Muzea vědy Buffalo New York, USA).
Nejnověji razí myšlenku přepravy na rostlinných kobercích i evolucionisté, kteří tak vysvětlují přítomnost kuskusovitých savců Ailurops ursinus a Strigocuscus celebensis na ostrově Celebes22 a lemurů na ostrově Madagaskar23. Roku 1995 podal jeden rybář zprávu o kolonizaci ostrova Anguilla v Západní Indii leguány. Ty vyhodily vlny na jednu z východních pláží ostrova, když je tam dopravil koberec z klád a vyvrácených stromů pár týdnů poté, kdy se dva hurikány přehnaly nad ostrovy Malých Antil. Vědci propočítali, že leguáni na koberci uplavali 320 km z Guadeloupe24.
Příznačné je, že biogeografové někdy vymezují oceány spíše než kontinenty jako hlavní biogeografické oblasti. Je tomu tak proto, že velmi často jsou patrné vzorce nespojitosti tam, kde je mnoho suchozemských organizmů rozeseto podél pobřeží nějakého oceánu. Biogeografovi z dvacátého století Léonu Croizatovi to bylo tak nápadné, že strávil mnoho času zakreslováním „tras“, aby tak zmapoval opakované výskyty těchto vzorců (25). Tam, kde se objevuje znovu určitá trasa použitá odlišnými organizmy skupinu po skupině, mluvíme často o „generalizované trase“. Například trasu rostliny Oreobolus znázorněnou na obrázku 3 sdílí spousta dalších rostlin a zvířat (26). Z těchto generalizovaných tras vyabstrahoval Croizat pět biogeografických „uzlů“ či „bran“, kudy se šíří rostliny a zvířata po světě (obrázek 4)27.
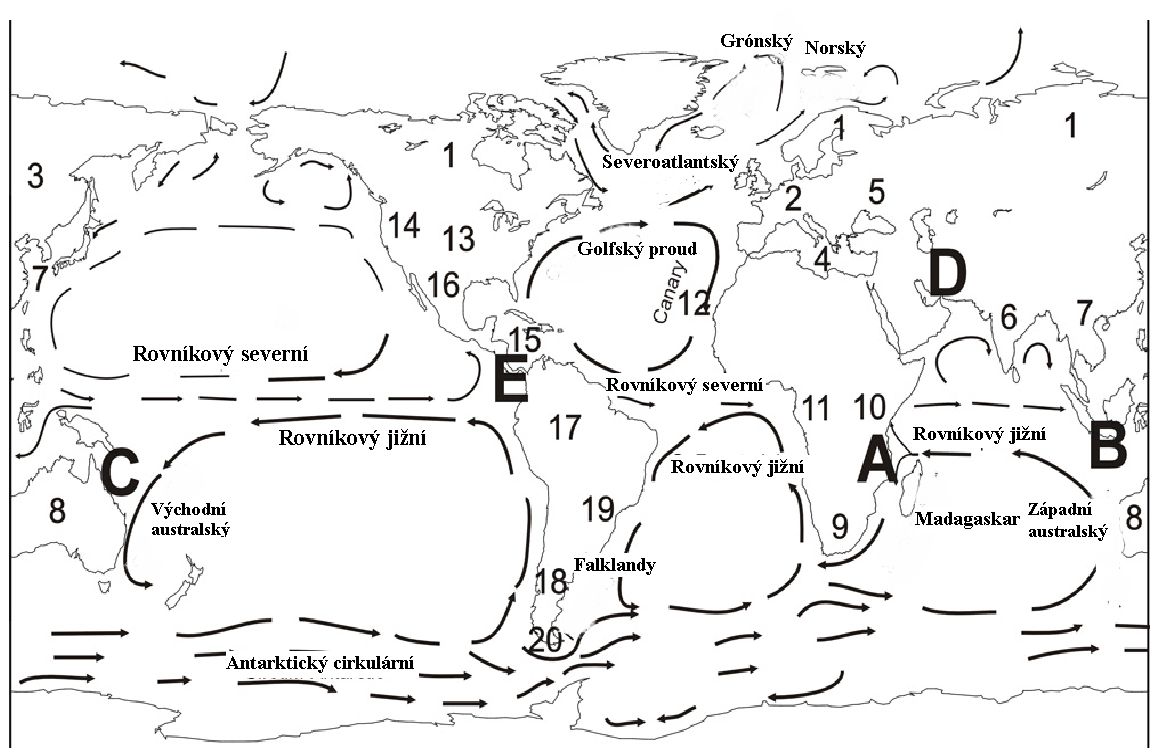
Obrázek 4. Přehled proudů, bran a oblastí endemizmu. Dvacet oblastí endemizmu vymezených de Candollem je označeno čísly 1 až 20. Pět biogeografických „bran“ vymezených Croizatem je označeno písmeny A až E. (Z Wise a Croxton, bibliogr. odkaz 30).
Zničující síla velkých objemů rychle tekoucí vody je nesmírná, a tak v raných stadiích Potopy podle Genesis zřejmě dokázala vyrvat ze země velké plochy lesů. Třebaže byla část z nich patrně pohřbena v usazeninách, mnoho miliard stromů asi unášela nadále voda jako obrovské „koberce z kmenů“28. Na těchto ostrovech vegetace pravidelně zavlažovaných deštěm se mohly snadno po delší čas udržet při životě rostliny i zvířata. Oceánské proudy pak dopravovaly tyto masivní „rafty“ po celé zemi; občas přitom přirazily ke břehu, kde mohla zvířata a hmyz „nastoupit“ či „vystoupit“, potom rafty opět proudy strhly do moře. Schopnost oceánských proudů přepravovat plovoucí předměty po světě jsme si ověřili nedávno, kdy roku 1992 nákladní loď ztratila v severním Tichém oceánu tisíce gumových kachniček, které se dávají dětem na hraní do vany. Za méně než dvacet let dopluly tyto hračky do Austrálie a Jižní Ameriky, a následně do Severního ledového oceánu i Atlantiku29. Teorii raftingu podporuje i zjištění profesorů Kurta Wiseho a Matthewa Croxtona, podle kterých výseky, které tvoří spojnice směru oceánských proudů s pevninami, odpovídají de Candollovým oblastem endemizmu i Croizatovým biogeografickým branám (obrázek 4)30. Tím nechceme říci, že suchozemští živočichové přežili Potopu na raftech, nýbrž že rafty jen usnadnily jejich rozšíření po zemi po Potopě, když se rozmnožili a opustili archu, když spočinula na horách Ararat (Genesis 8:4).
Transport prostřednictvím lidí
Podle Bible se lidstvo po zmatení jazyků v Bábelu (Genesis 11) rozptýlilo po celé zemi31. Závažné podpůrné důkazy pro tento fakt nalézáme v archeologii, podobnostech jazyků, kterými mluví lidé v Evropě i na Dálném východě, i v anatomických analýzách a analýzách DNA32. Je tedy nasnadě představa, že mnozí z těchto lidí cestujících do různých krajů zřejmě s sebou vzali zvířata jako potravu na cestu i za účelem pěstování zemědělství po příchodu do místa určení33.
Migrace a částečné vymření
Mnoho kreacionistů má za to, že brzy po Potopě35 nastala doba ledová34. Během ní patrně klesla hladina moří, jelikož se voda nahromadila v pevninských ledových příkrovech, a mohly se také vytvořit ledové mosty mezi světadíly, přes které mohla migrovat zvířata. Většina evolucionistů věří, že kdysi existoval takový most přes Beringovu úžinu spojující Asii s Amerikou36. Mnoho geologů si myslí, že po oddělení světadílů došlo k mohutným tektonickým zdvihům37 a mosty mezi kontinenty, které kdysi existovaly v jiných částech světa, se pak zřejmě dostaly pod hladinu moře. Zvířata mohla přecházet z jednoho světadílu na druhý přes tyto mosty poté, co se rozmnožila a rozprchla z archy, možná po stovky let. Rychlost, kterou mohou zvířata takto osídlit rozsáhlá území, lze doložit na australských králících. Před příchodem Evropanů na tomto světadílu králíci nežili, ale roku 1859 byla jedna jejich kolonie vysazena v jižní Viktorii, na jihovýchodě. Během padesáti let pak králíci osídlili celý světadíl až po západní pobřeží38.
Je jisté, že na jednotlivých světadílech docházelo k velkým klimatickým změnám. Mamuti, nosorožci, bizoni, koně i antilopy například kdysi žili ve velkém množství na severní Sibiři. Egyptské pouště byly kdysi bujnými savanami39. Skupiny zvířat, kterým se kdysi dobře dařilo v jistých oblastech, mohly na těchto místech vymřít, a přežili zřejmě jen ti jedinci, kteří se odstěhovali na jiné světadíly. Ba co víc, klimatické změny a konkurence jiných zvířat mohly být docela dobře impulzem k migraci. Na druhé straně můžeme nepřítomnost konkrétních skupin na konkrétních světadílech chápat jako důkaz toho, že se tam nikdy neodstěhovala ani tam nebyla dopravena, takže tam nepřežila.
Speciace
Na rozdíl od častých výroků těch, kdo se snaží zdiskreditovat kreacionizmus, většina kreacionistů netvrdí, že druhy jsou neměnné. Naopak, argumentují silně ve prospěch procesu speciace. Kromě toho, že pro speciaci svědčí pádné vědecké důkazy, je tento proces také stěžejní složkou biblického výkladu rozmanitosti života pozorovaného dnes na zemi. Podle Bible byla jedinými suchozemskými zvířaty, která přežila Potopu, ta, která zachránil Noe. Na palubě archy byly zastoupeny všechny druhy zvířat a jejich potomky musí být tedy všechny nyní žijící zoologické druhy (Genesis 6-8). Bibličtí kreacionisté v zásadě věří, že genetickou informaci potřebnou pro vznik všech zoologických druhů nesla zvířata, která se vylodila z archy. Měli bychom však zopakovat, že bibličtí kreacionisté nevěří, že se při speciaci mohou křížit druhy, takže se například plaz nikdy „speciací“ nezmění v savce ani lidoop v člověka.
Jelikož kreacionisté připouštějí, že zvířata i rostliny byly stvořeny se schopností přizpůsobit se novým biotopům, zastávají názor, že přítomnost podobných zoologických druhů či rodů v sousedících biotopech může být někdy vysvětlena biologickou proměnou.
Závěr
I když biogeografická pozorování přinášejí silné důkazy pro proces speciace, nepodporují obecnější předpovědi evoluční teorie ani model pomalého kontinentálního driftu zastávaný geology staré země. Data lze však interpretovat jako potvrzení biblické zprávy o rekolonizaci a rozrůznění organizmů po Potopě podle Genesis.
Odkazy
1. Wood, T.C., A Creationist Review and Preliminary Analysis of the History, Geology, Climate and Biology of the Galápagos Islands, Eugene, Wipf and Stock, OR, pp. 108–125, 2005.
2. Wood, T.C. and Murray, M.J., Understanding the Pattern of Life, Broadman & Holman, Nashville, TN, p. 192, 2003.
3. Humphries, C. and Parenti, L., Cladistic Biogeography: Interpreting Patterns of Plant and Animal Distributions, 2nd edn., Oxford University Press, Oxford, pp. 21–22, 1999.
4. Nelson, G. and Platnick, N., Systematics and Biogeography: Cladistics and Vicariance, Columbia University Press, New York, pp. 368, 524, 1981; Cox, C.B., The biogeographic regions reconsidered, Journal of Biogeography 28:511–523, 2001; www.interscience.wiley.com.
5. Ridley, M., Evolution, 3rd edn., Blackwell Science, Oxford, ch. 17, 2004.
6. George, W. and Lavocat, R., The Africa–South America Connection, Clarendon Press, Oxford, p. 159, 1993; Davis, C. et al., High-latitude tertiary migrations of an exclusively tropical clade: evidence from Malpighiaceae, International Journal of Plant Sciences 165 (4 Suppl.):S107–S121, 2004; www.people.fas.harvard.edu/~ccdavis/pdfs/Davis_et_al_IJPS_2004.pdf.
7. George and Lavocat, ref. 6, ch. 9.
8. George and Lavocat, ref. 6, p. 159.
9. Thorne, R., Major disjunctions in the geographic ranges of seed plants, The Quarterly Review of Biology 47(4):365–411, 1972; Buffalo Museum of Science (New York), Panbiogeography: Pacific Basin Tracks, sciencebuff.org/pacific_basin_tracks.php.
10. Allaby, M., Dromiciopsia; in: A Dictionary of Zoology, Oxford University Press, Oxford, 1999; www.encyclopedia.com.
11. Beck, W. et al., Life. An Introduction to Biology, 3rd edn., HarperCollins, New York, p. 1324, 1991.
12. Schatz, G., Malagasy/Indo-Australo-Malesian phytogeographic connections; in: Lourenço, W.R. (Ed.), Biogeography of Madagascar, Editions Orstom, Paris, 1996, www.mobot.org.
13. Qian, H., Floristic relationships between eastern Asia and North America: test of Gray’s hypothesis, The American Naturalist 160(3):317–332, 2002.
14. Smiley, C., Pre-Tertiary phytogeography and continental drift: some apparent discrepancies; in: Gray J. and Boucot, A. (Eds.), Historical Biogeography, Plate Tectonics and the Changing Environment, Oregon State University Press, Corvallis, OR, pp. 311–319, 1976.
15. Cifelli R. and Davis, B., Marsupial origins, Science 302:1899–1900, 2003.
16. Quantum, ABC, 6 November 1991, cited in Focus: news of interest about creation and evolution, Creation 14(2):5–8, 1992; Gish, D., Evolution: The Fossils Still Say No! Institute for Creation Research, El Cajon, CA, pp. 178–183, 1995.
17. Flannery, T., Forum: A hostile land—could one tiny fossil overthrow Australia’s orthodoxy? New Scientist 2116:47, 1998.
18. Simpson, G.G., Mammals and land bridges, Journal of the Washington Academy of Sciences 30:137–163, 1940; www.wku.edu; Thorne, R., Major disjunctions in the geographic ranges of seed plants, The Quarterly Review of Biology 47(4):375, 1972; Hagen, A., Trans-Atlantic dispersal and phylogeography of cerastium arcticum (Caryophyllaceae) inferred from RAPD and SCAR markers, American Journal of Botany 88(1):103–112, 2001.
19. Another remarkable example of “convergent evolution” is the echolocation systems used by bats and whales, which, again, are believed to have evolved quite separately. That such similar and very sophisticated systems could have arisen through chance mutations stretches credibility a very long way; Spetner, L., Not by Chance, Judaica Press, New York, 1998.
20. Nelson,G. and Platnick, N., Systematics and Biogeography: Cladistics and Vicariance, Columbia University Press, New York, p. 223, 1981.
21. Moody, P., Introduction to Evolution, Harper & Brothers, New York, p. 262, 1953.
22. Heinsohn, T., A giant among possums, Nature Australia 26(12):24–31, 2001.
23. Tattersall, I., Madagascar’s lemurs, Scientific American 268(1):110–117, January 1993; p. 90.
24. Censky, E. et al., Over-water dispersal of lizards due to hurricanes, Nature 395:556, 1998; Yoon, C., Hapless iguanas float away and voyage grips biologists, The New York Times, 13 March 2008; www.nytimes.com.
25. Humphries. C. and Parenti, L., Cladistic Biogeography: Interpreting Patterns of Plant and Animal Distributions, 2nd edn., Oxford University Press, Oxford, pp. 33, 37, 1999; Croizat, L., Panbiogeography, vols. 1, 2A and 2B, self-published, 1958.
26. Seberg, O., Taxonomy, phylogeny, and biogeography of the genus Oreobolus R.Br. (Cyperaceae), with comments on the biogeography of the South Pacific Continents, Botanical Journal of the Linnean Society 96:119–195, 1998; Buffalo Museum of Science (New York), Vicariance biogeography and panbiogeography of the plant genus Oreobolus (Cyperaceae): a comparison of methods and results; www.sciencebuff.org/panbiogeography_of_oreobolus.php.
27. Croizat, L., Panbiogeography, vol. 2B, fig. 259, p. 1018.
28. Scheven, J., The Carboniferous floating forest an extinct pre-Flood ecosystem, Journal of Creation (CEN TJ) 10(1):70–81, 1996; Wieland, C., Forests that grew on water: startling hard facts from coal uproot the millions of years idea, Creation 18(1):20–24, December 1995.
29. Ford, P., Drifting rubber duckies chart oceans of plastic, Christian Science Monitor, 31 July 2003, www.csmonitor.com; Clerkin, B., Thousands of Rubber Ducks to Land on British shores after 15 year journey’, Daily Mail, 27 June 2007, www.dailymail.co.uk.
30. Wise, K.P. and Croxton, M., Rafting: a post-Flood biogeographic dispersal mechanism, Proceedings of the Fifth International Conference on Creationism, Creation Science Fellowship, Pittsburgh, PA, pp. 465–477, 2003.
31. In this, the Bible is emphatic. According to Genesis 11:8, “ … the LORD scattered them from there over all the earth”; and again, in Genesis 11:9, “From there the LORD scattered them over the face of the whole earth” (my italics).
32. Schroeder, L.L., A possible post-Flood human migration route, Journal of Creation (TJ) 19(1):65–72, 2005.
33. Woodmorappe, J., Causes for the biogeographic distribution of land vertebrates after the Flood, Proceedings of the Second International Conference on Creationism, Pittsburgh, PA, pp. 361–370, 1990.
34. That is, they believe in one Ice Age, from around 2350 to 1500 BC, See Garner, P., The New Creationism, Evangelical Press, Darlington, UK, ch. 15, 2009.
35. Batten, D. (Ed.), The Creation Answers Book, Creation Book Publishers, ch. 16, 2009.
36. Elias, S. et al., Life and times of the Bering Land Bridge, Nature 382:60–63, 1996.
37. Many biblical creationists also believe that the continents separated, but not over millions of years. They understand this to have happened very rapidly, during or soon after the Genesis Flood. Dr John Baumgardner of the Los Alamos National Laboratory, USA, has produced a computer model of the earth’s mantle, and has shown that tectonic plate movement could have caused the continents to separate both spontaneously and very rapidly. The model also proposes a possible mechanism for the global Flood (Batten, D. (Ed.), The Creation Answers Book, Creation Book Publishers, 157–166, 2009.
38. Wieland, C., The grey blanket, Creation 25(4):45–47, 2003.
39. Fitzpatrick, T., Scientists find fossil proof of Egypt’s ancient climate, Washington University, St Louis; www.wustl.edu.
Tento článek je též v ruštině